



铝单板厂家电话价格优惠现货促销(新抗原疗法)
发布时间:2023-04-07 08:30:48 人气:25 来源:铝单板厂家
转自:药时空继2022年11月的Non-viral precision T cell receptor replacement for personalized cell therapy个性化的neoTCRs之后,2023年3月发表了Neoantigen-targeted CD8+ T cell responses with PD-1 blockade therapy研究了抗PD-1免疫治疗对其neoTCR-T细胞的应答反应。
新抗原是由HLAs呈现的非同义突变产生的多肽,可被抗肿瘤T细胞识别HLA等位基因多样性和有限的临床样本限制了对患者治疗过程中新抗原靶向T细胞反应的研究在这里,应用最近开发的技术从血液和肿瘤中捕获转移性黑色素瘤患者的新抗原特异性T细胞,无论是否对anti-PD-1免疫治疗有反应。
建立了个性化的新抗原-HLA捕获文库,用于单细胞分离T细胞并克隆其T细胞受体(neoTCRs)具有不同neoTCR序列(T细胞克隆型)的多个T细胞在来自7名具有长期临床反应的患者的样本中识别出有限数量的突变。
随着时间的推移,这些neoTCR克隆型在血液和肿瘤中被反复检测到来自4名对抗PD-1无反应的患者的样本也显示血液和肿瘤中的新抗原特异性T细胞对数量有限的TCR多克隆较低的突变有反应,并且在连续样本中没有反复检测到。
使用非病毒CRISPR-Cas9基因编辑重建供体T细胞中的neoTCR,显示了对患者匹配的黑色素瘤细胞系的特异性识别和细胞毒性因此,有效的抗PD-1免疫治疗与肿瘤和血液中存在针对有限数量的免疫显性突变的多克隆CD8+T细胞有关,这些突变随着时间的推移而被反复识别。
推测对免疫检查点阻断(ICB)治疗诱导的突变新抗原特异性T细胞反应谱分析需要①一种高灵敏度的方法,可以在相对较小的患者来源的样本中检测T细胞对由不同的HLA呈现的数百种假定的新抗原的反应,其中包含从肿瘤活检组织中扩增的几百万个PBMC或TIL;
②在时间有效的过程中重建分离的新抗原特异性TCRs(neoTCRs);以及③评估T细胞对内源性表达突变新抗原的匹配患者自体细胞株的识别和抗肿瘤活性从11例接受PD-1阻断免疫治疗的转移性黑色素瘤患者中收集了不同时间点的PBMC,并从可用的患者肿瘤活检组织中建立了TIL培养物和自体肿瘤细胞系。
对肿瘤细胞系或肿瘤活检组织进行全外显子组测序(WES)和RNA测序(RNA-seq)分析,并与从匹配患者的PBMC中获得的正常对照DNA进行比较这些数据被用来定义每个患者的6个HLAI类等位基因,检测患者特有的非同义突变,并对患者的任何HLAI类分子的任何一个可能呈现的新抗原进行计算预测。
根据肿瘤的表达水平和预测的与患者自身的HLAI类分子的结合亲和力,对预测的新抗原进行优先排序优先列出了每个患者多达243个(中位数172个,范围17-243)新抗原肽-HLA候选列表(肽长度,8-11个aa),覆盖在多达64个HLAI类等位基因的HLA文库中。
在Expi293F细胞中以单链三聚体的形式构建多肽-HLA复合物文库,将新抗原肽基因序列依次融合到β-2-微球蛋白(β2M)结构域和HLA然后,这些个性化的多肽文库被DNA条形码、荧光标记和多聚体用作捕捉试剂。
一旦组装完成,这些文库就与患者的PBMC或从肿瘤活检组织中扩增的淋巴细胞孵育,并通过单细胞分选获得新抗原特异性T细胞值得注意的是,测序数据和所有TIL培养物都是从每个患者的相同病变的活检组织中获得的T细胞分离包括CD39、CD103和CD95等抗原敏感的T细胞标志物。
每个预测的新抗原-HLA多聚体特有的条形码能够破译捕获的T细胞的抗原特异性(图1a),因此定义了抗肿瘤CD8+T细胞识别的突变新抗原。
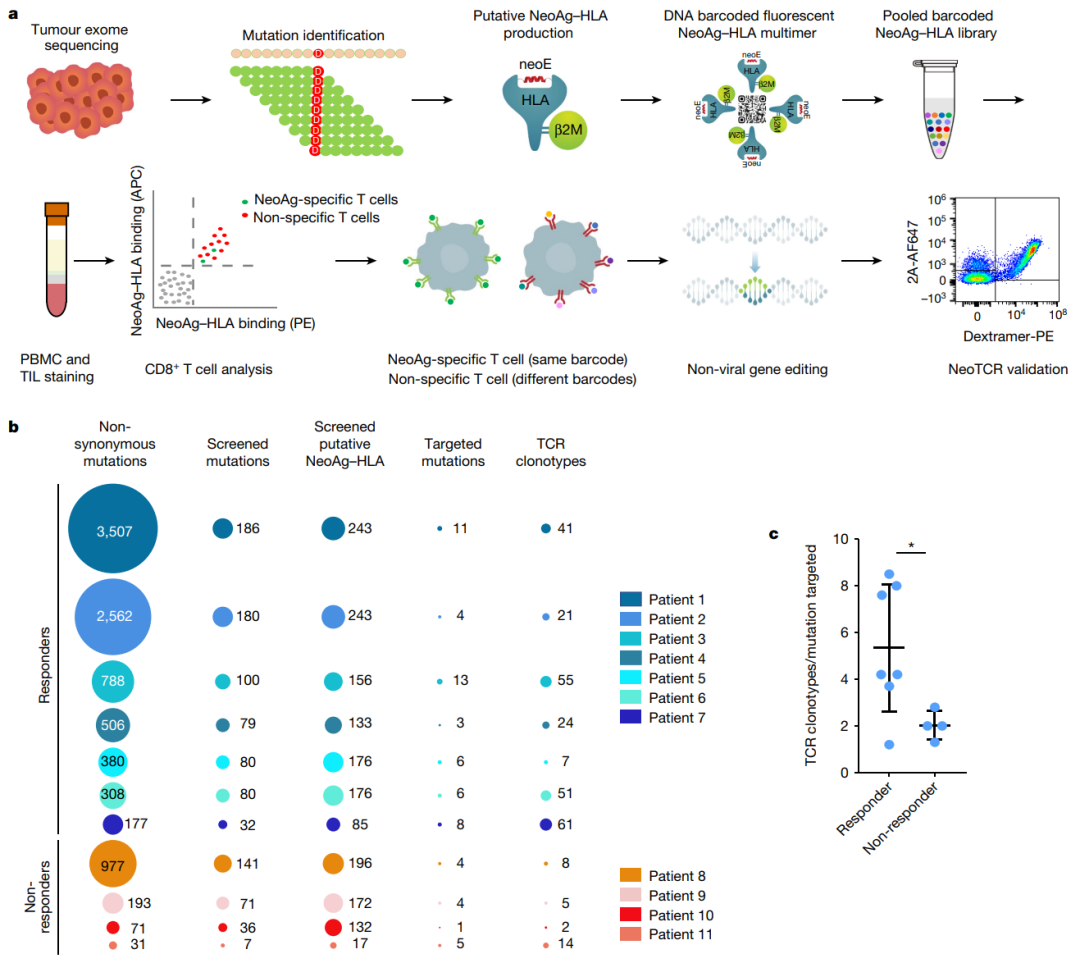
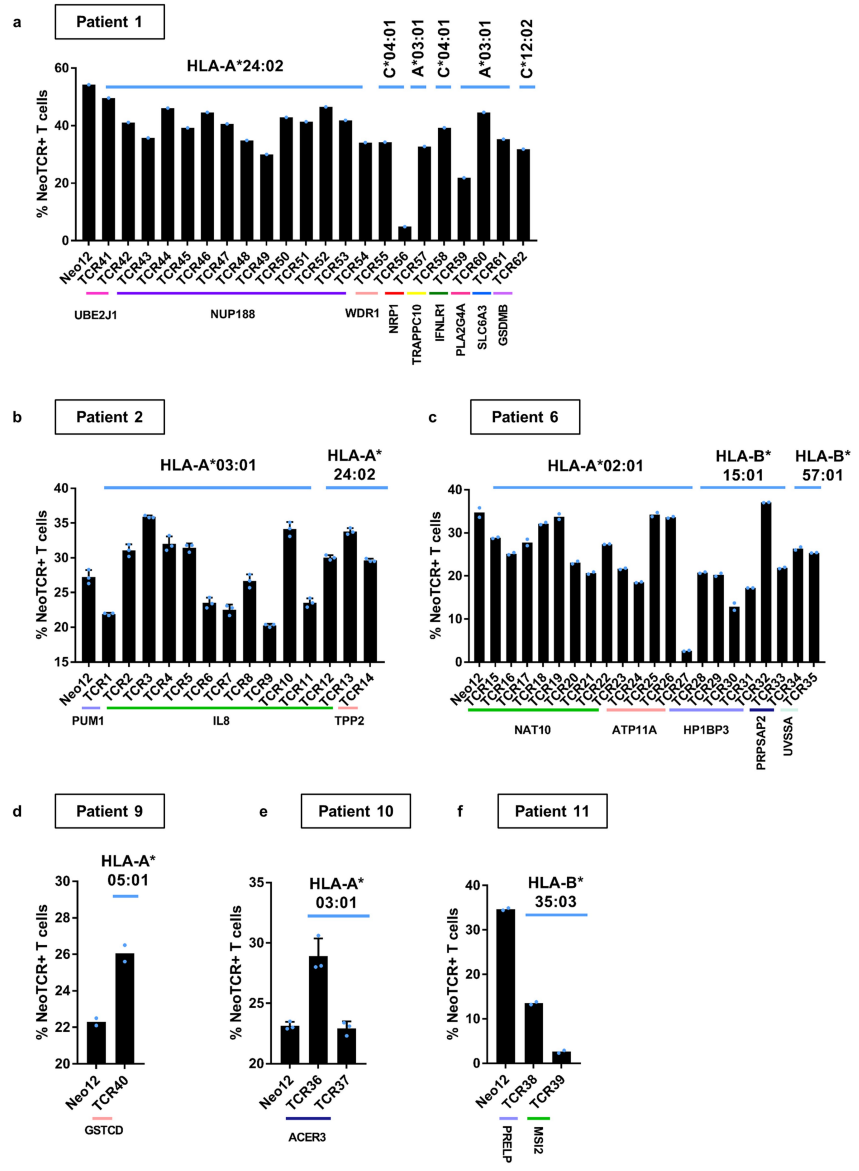
●图1 新抗原特异性T细胞筛选和TCR克隆型鉴定a:从患者样本中分离新抗原特异性TCR的示意图b:在对抗PD-1治疗有反应或无反应的患者(患者1-7)或无患者(患者8-11)中分离的非同义突变、筛选的突变、预测的新抗原-HLA复合物、新抗原特异性T细胞靶向突变和新抗原特异性T细胞克隆型的数量。
c:每个患者中靶向每个突变分离出的新抗原特异性TCR克隆型的比率对11名转移性黑色素瘤患者的样本进行了这一过程,其中7人对抗PD-1治疗有临床反应,3人没有反应,1人在收集基线样本后和接受第一次计划的Nivolumab输注之前迅速进展
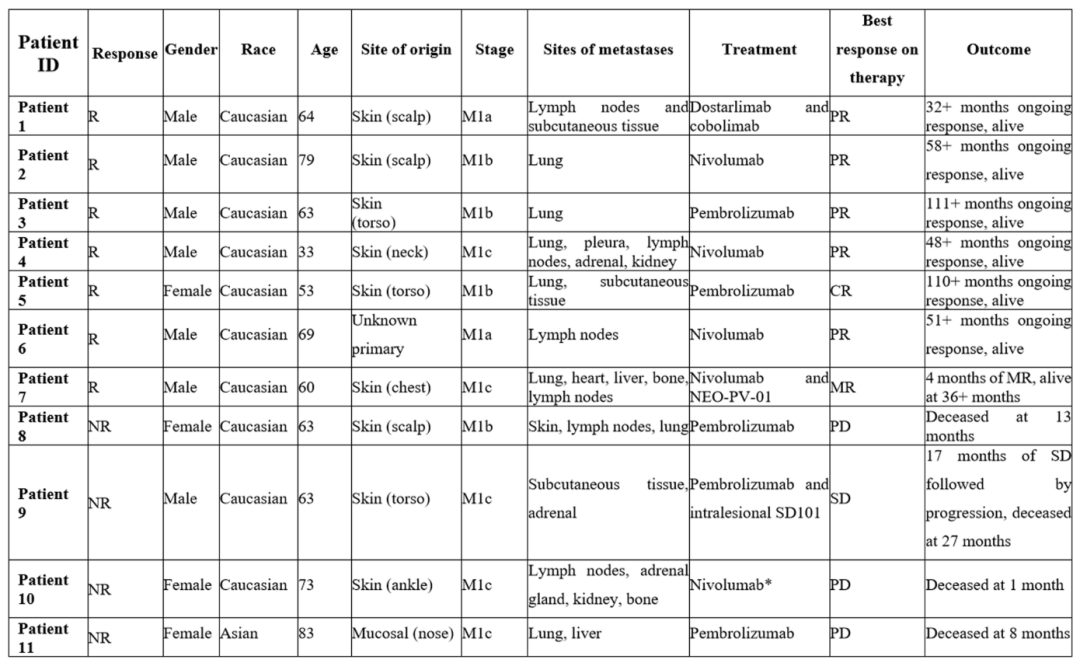
(表1)在报告时,有临床反应的7名患者仍活着患者1对Dostarlimab(抗PD-1抗体)和cobolimab(抗TIM3抗体)有长期的临床反应(持续32个月以上),患者2至6对使用抗PD-1抗体Nivolumab或Pembrolizumab的单药治疗有长期的临床反应(持续48至111个月)。
患者7开始治疗的是一个较大的转移性黑色素瘤,转移到五个器官(包括心脏和肝脏),几个转移灶的大小缩小,并伴随着其他部位的进展,这代表着自治疗开始以来持续了36个多月的混合临床反应4名没有临床反应的患者在报告时已经死亡。
患者8单剂Pembrolizumab后病情恶化患者9接受Pembrolizumab联合肿瘤内注射Toll样受体9激动剂SD101,导致注射的皮下部位消退但远端肾上腺转移无反应,病情稳定17个月随后广泛的骨和腹膜转移。
患者10在计划的首次注射Nivolumab之前进展迅速患者11在接受Pembrolizumab治疗后4个月内进展(表1)。除了患者11黏膜黑色素瘤外,所有患者的起源都是角质黑色素瘤。表1 患者特征
对患者来源的黑色素瘤细胞株和活检组织的WES分析发现了广泛的突变,从患者11中的31个到患者1中的3507个(中位数,380)抗PD-1治疗有反应的患者的黑色素瘤的肿瘤突变负荷中位数(中位数,506个非同义突变)高于治疗无效的患者的样本(中位数,132个非同义突变)。
尽管表达的突变范围很广,但被T细胞识别为新抗原的突变数量变化要小得多,范围从1到13(图1b,图E1和表S1)因此,有证据表明免疫优势即T细胞对突变的新抗原的反应与突变的数量不是线性相关的,而是针对一组有限的突变,这些突变在对抗PD-1治疗有临床反应和没有临床反应的患者中被识别为新抗原。
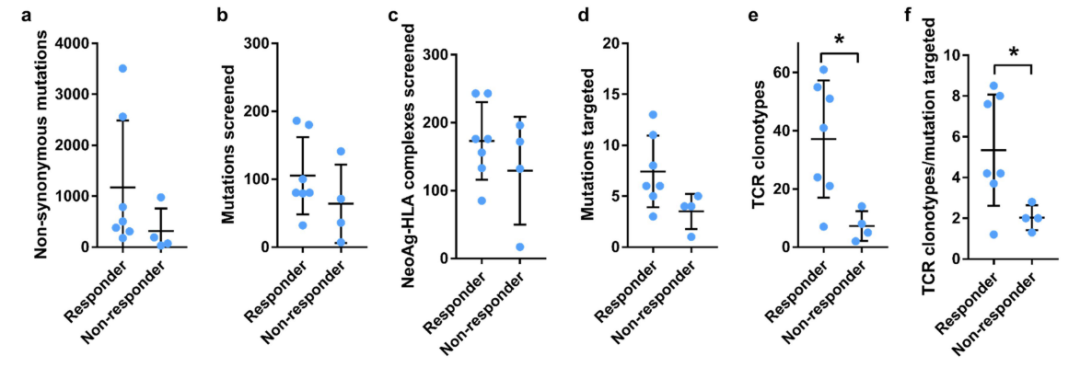
●图E1 新抗原特异性T细胞分离及TCR克隆型鉴定。对治疗有反应和无反应患者的新抗原特异性T细胞进行纵向横向分析,总结关键参数。表S1 患者的突变与靶向新抗原
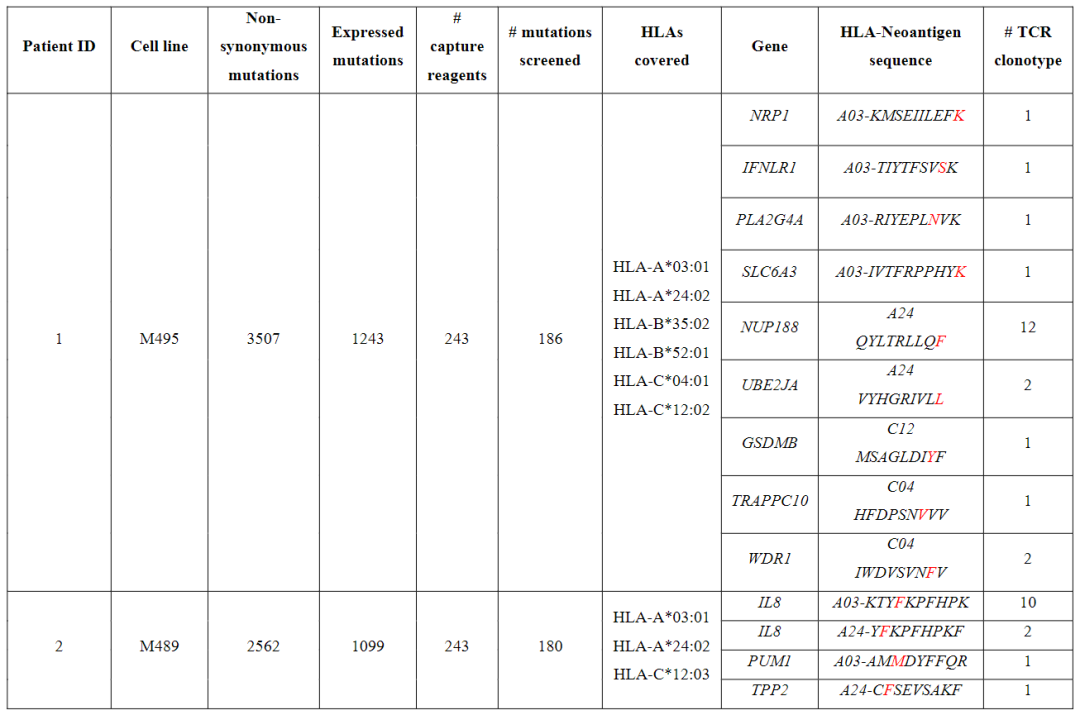
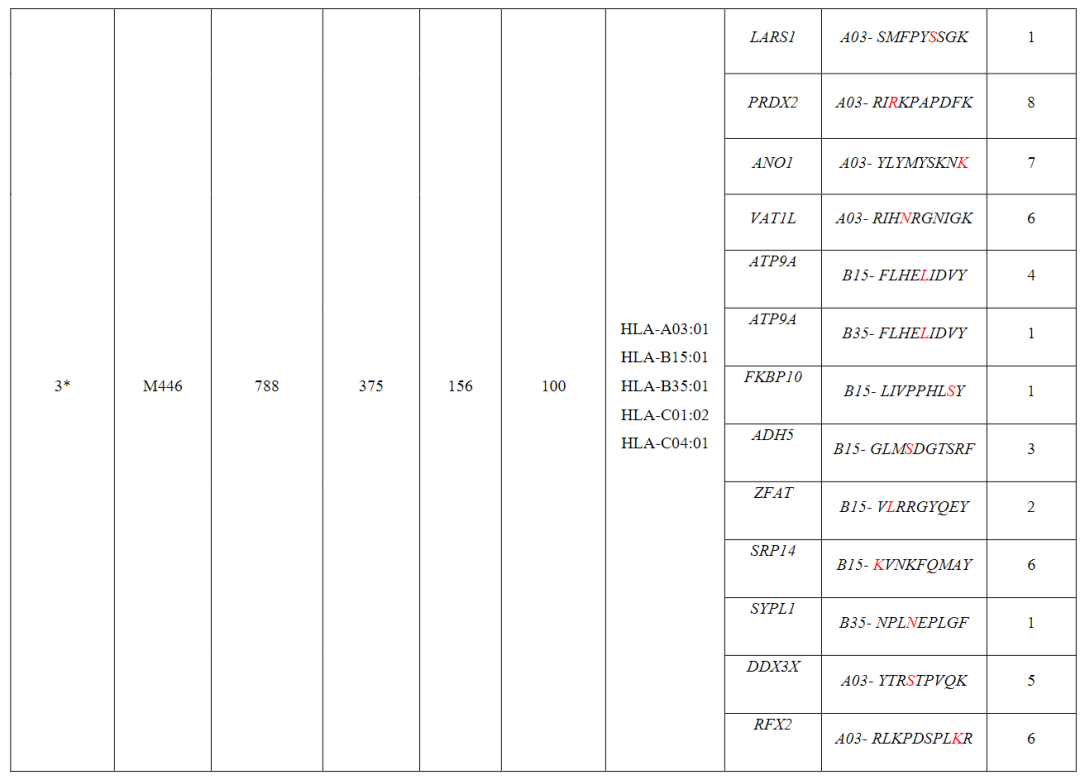
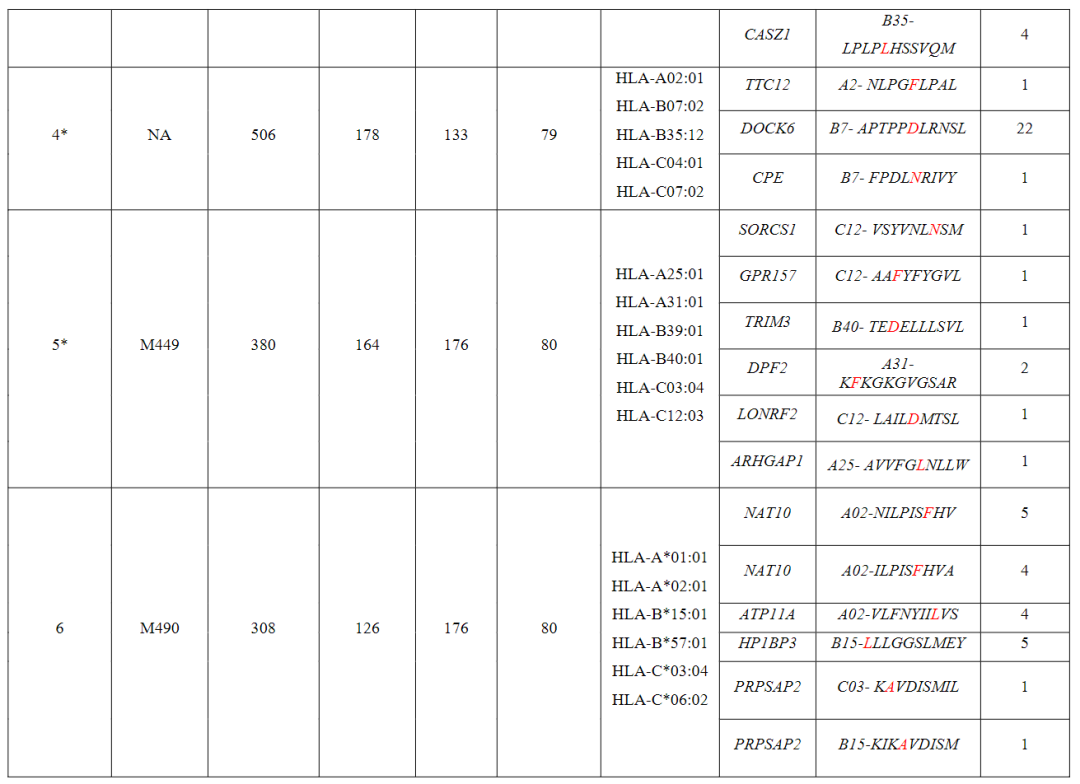
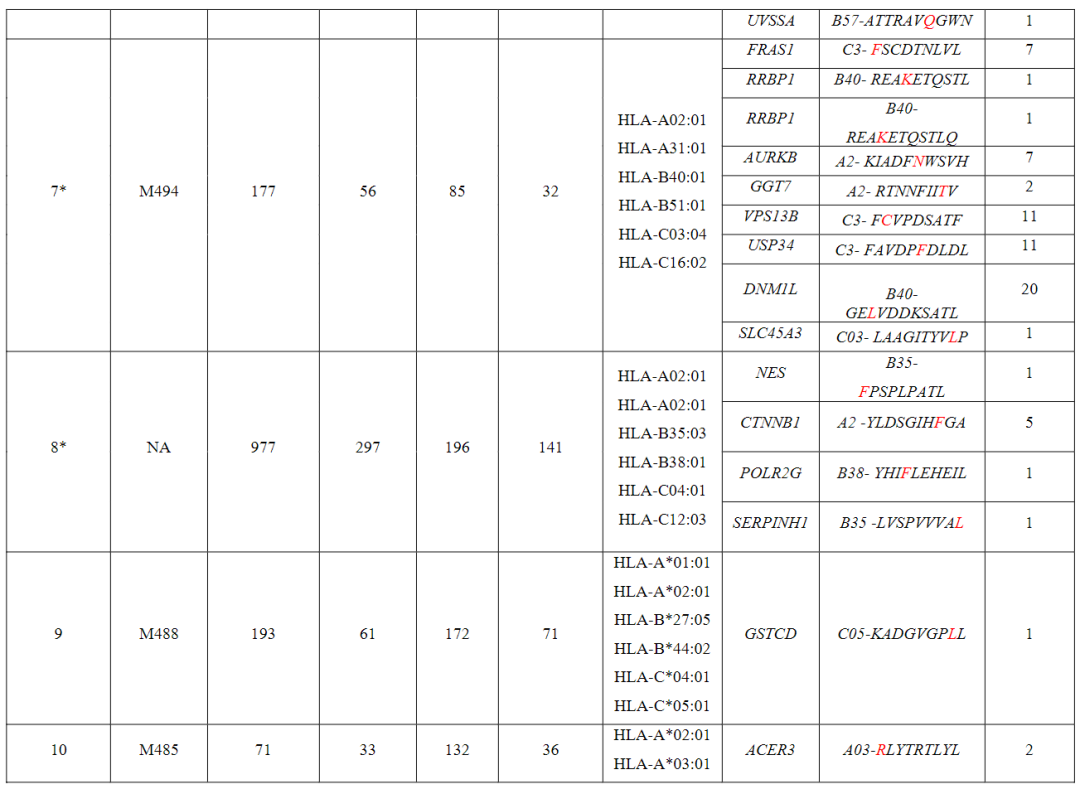
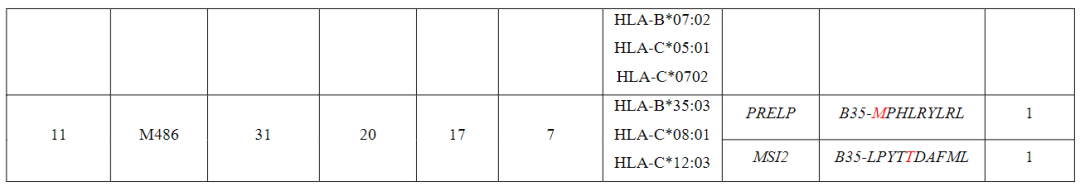
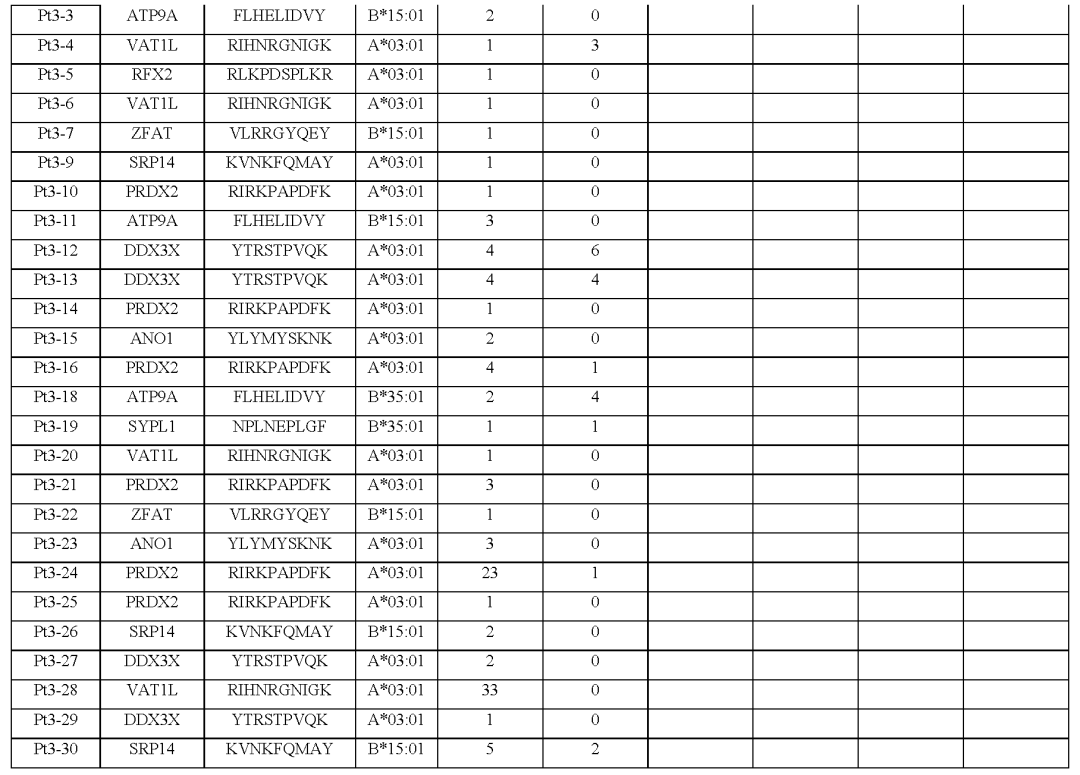
从对治疗有临床反应的患者(患者1至7)的血液和肿瘤中分离出的新抗原特异性T细胞的单细胞TCR测序显示,每个突变靶向中位数为5个(范围为1至23)单个TCR-α和β链对,每个患者中位数为41个(范围为7至61个)分离的neoTCR克隆型。
相比之下,4名对治疗没有临床反应的患者(患者8至11)显示,每个突变靶向中位数为2个(范围1至6)neoTCR克隆型,每个患者中位数为6.5(范围2至14)分离的neoTCR克隆型(图1b,c,图E1和表S1)
接下来,对每个患者的所有可用顺序PBMC和TIL样本进行了新抗原识别的纵向分析患者1在头皮的淋巴结和皮下组织有持久的反应(图2a,b)在来自患者1的黑色素瘤细胞系中,总共发现了3507个体细胞编码突变,其中1243个被RNA-seq预测表达(表S1)。
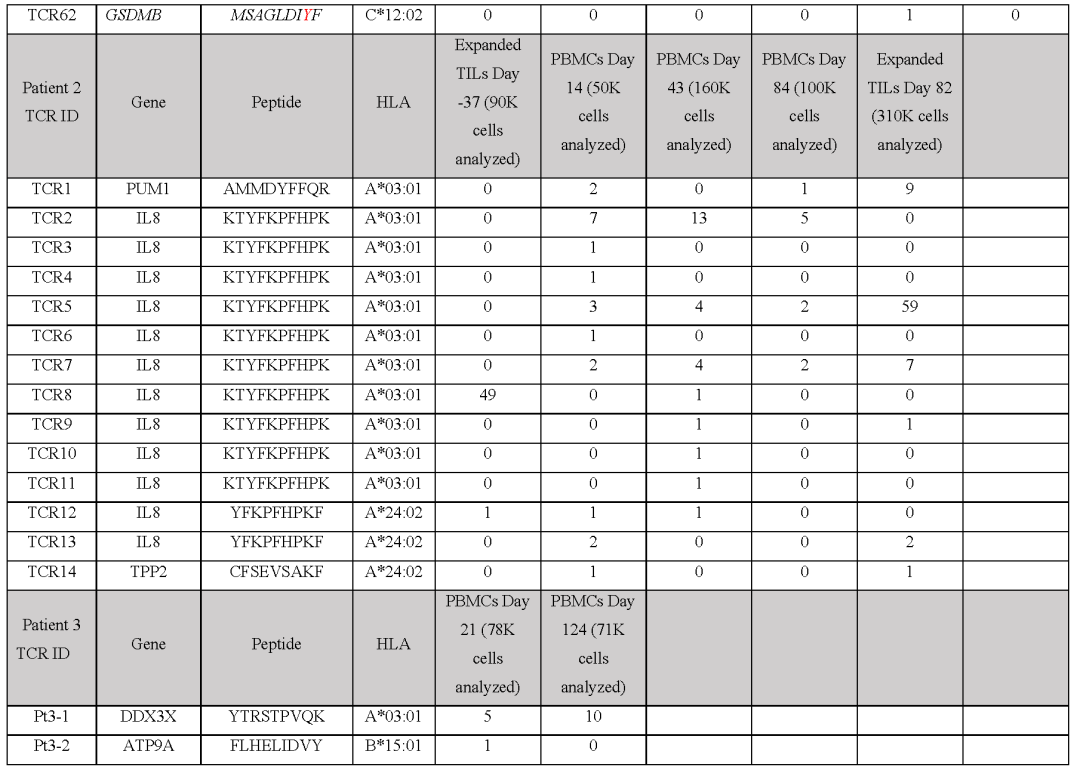
建立了一个由243种新抗原-HLA捕获试剂组成的文库,涵盖了患者6个HLAI类呈现的186个突变,并用于从血液和肿瘤样本中筛选CD8+T细胞(图2c和表S1)共分离出571个新抗原特异性T细胞,包括41个不同的neoTCR克隆型和11个靶向突变。
在分离出新抗原特异性T细胞后,选择22个neoTCR进行TCR克隆、在健康供者T细胞中的表达以及新抗原特异性表位与HLA的结合验证这22个neoTCR存在于总共482个新抗原特异性T细胞中,并针对9个不同的突变。
在分析的不同时间点,NUP188、UBE2J1和WDR1的突变被认为是新抗原(图2c和表S2)在具有不同T细胞频率的连续血液和肿瘤样品中检测到22种不同的neoTCR克隆型,一些在不同的时间点扩增或收缩。
值得注意的是,外周抽血在5mL到20mL之间,肿瘤活检是核心针活检,在任何时间点只采集每个隔室中的少数T细胞类似的发现在患者2和6的样本中也很明显,他们分别对肺(图2d,e)和淋巴转移(图2g,h)有持久的抗PD-1治疗反应。
患者2的自体细胞系有2562个非同义突变,其中1099个表达患者特异性文库包含243种捕获试剂,涵盖患者三个HLA呈现的180个突变靶向4个突变,分离出21个不同的neoTCR克隆型共筛选出19个neoTCR克隆型用于TCR克隆、在健康供者T细胞中的表达和特异性新表位-HLA结合验证,其中14个获得验证。
总共分离的186个新抗原特异性T细胞(表S2)包括这14个不同的靶向3个突变的neoTCR克隆型中的一个(图2f,表S1和2,图E1)对于患者6,在308个体细胞编码突变中,有126个表达患者特异性文库包括176种捕获试剂,覆盖80个突变,由6名患者的HLAI类分子呈现。
总共分离出针对6个突变的51个不同的neoTCR克隆型其中27个克隆型被克隆并在健康供者T细胞中表达,其中21个克隆型经新表位-HLA结合验证这21个neoTCR克隆型在296个分离的新抗原特异性T细胞中被鉴定出来,并针对5个突变。
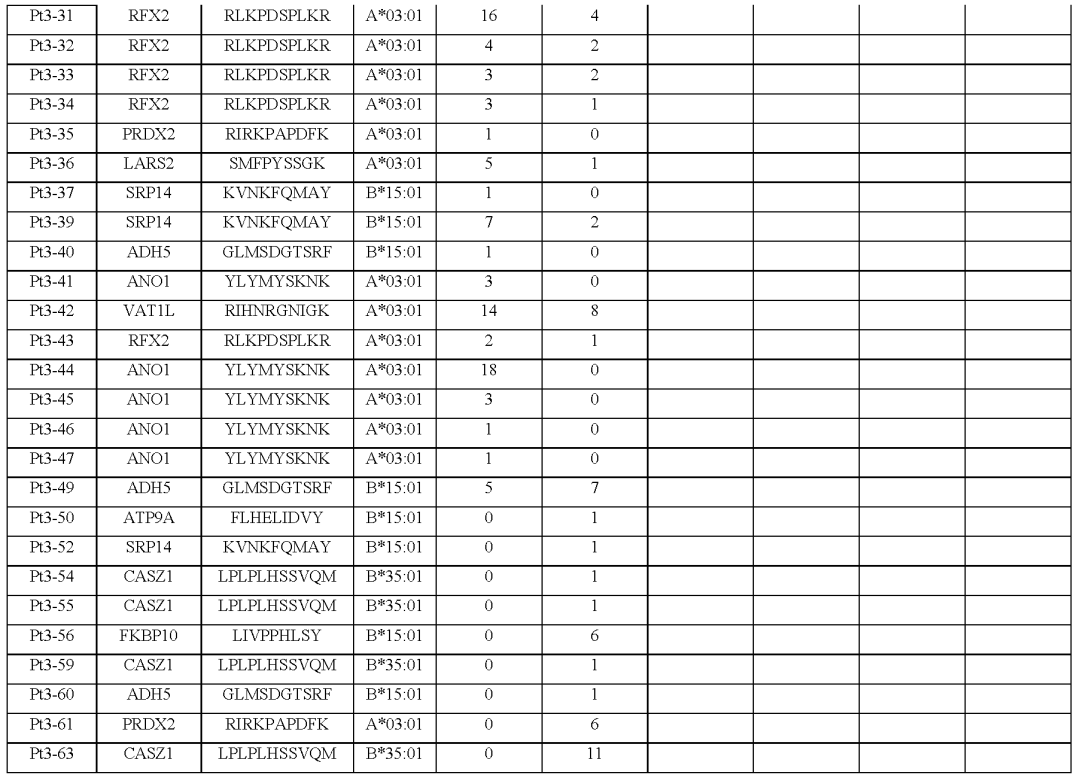
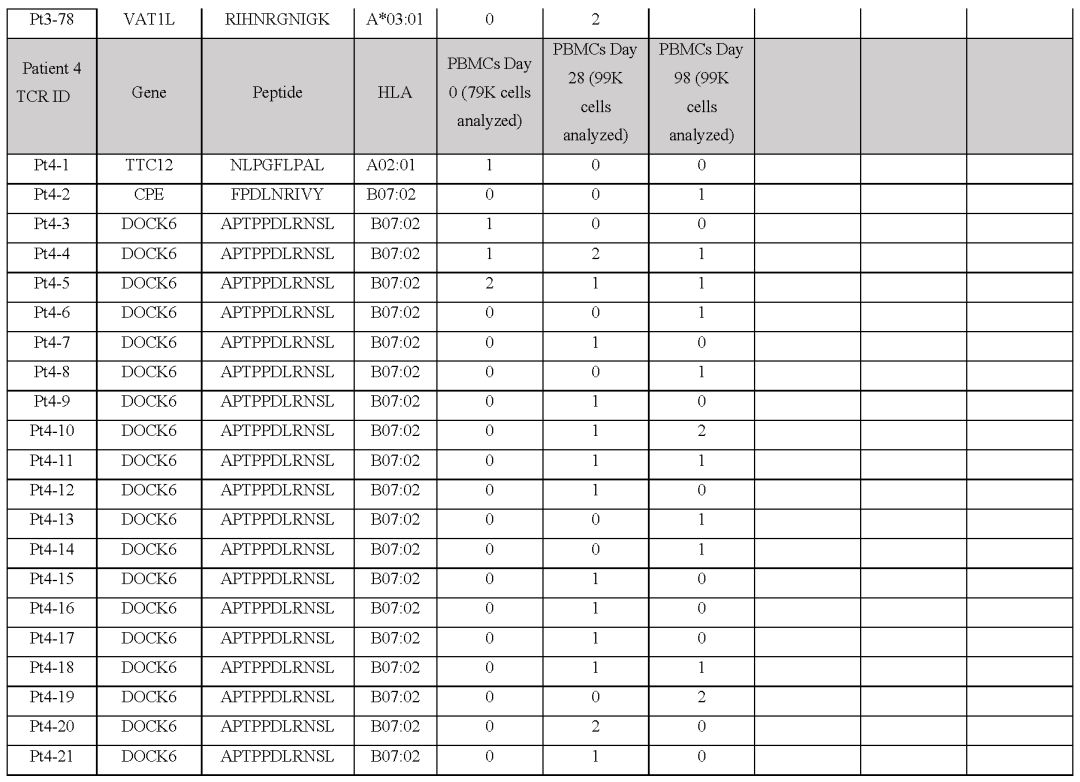
(图2i,表S1和2,图E1)对治疗有反应的患者3、4、5和7,自体细胞系呈现788(患者3)、380(患者5)和177(患者7)非同义突变,患者4的肿瘤活检组织显示506个体细胞编码突变在这些突变中,分别表达了375、178、164和56个突变。
患者特异性文库包括156、133、176和85种捕获试剂,分别涵盖100、79、80和32个突变从这些患者中分别分离出55、24、7和61个针对13、3、6和8个突变的neoTCR(图E1和E2以及表S1,S2)
。
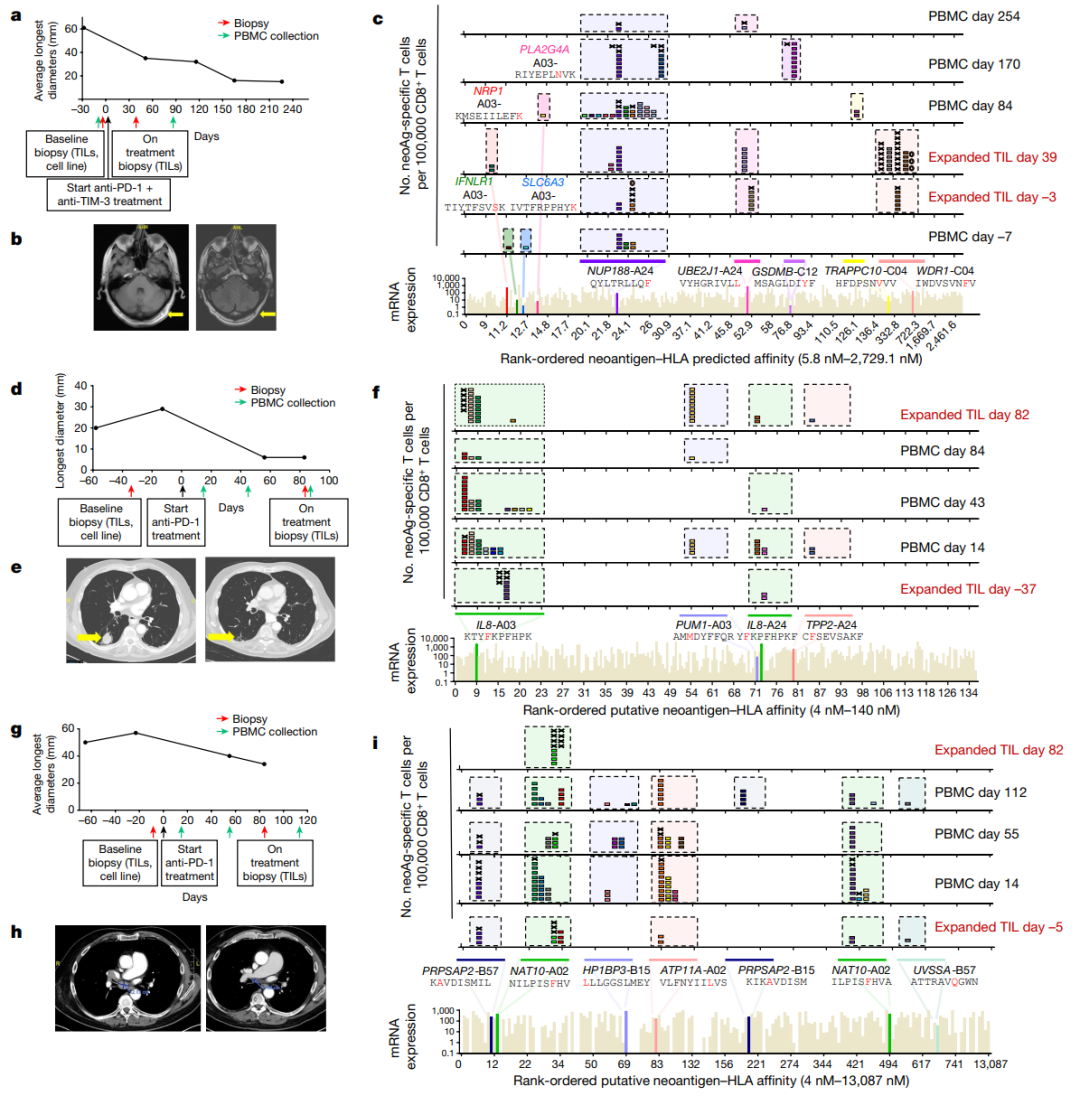
●图2 抗PD-1治疗后患者TIL和PBMCs中新抗原特异性T细胞的分离。
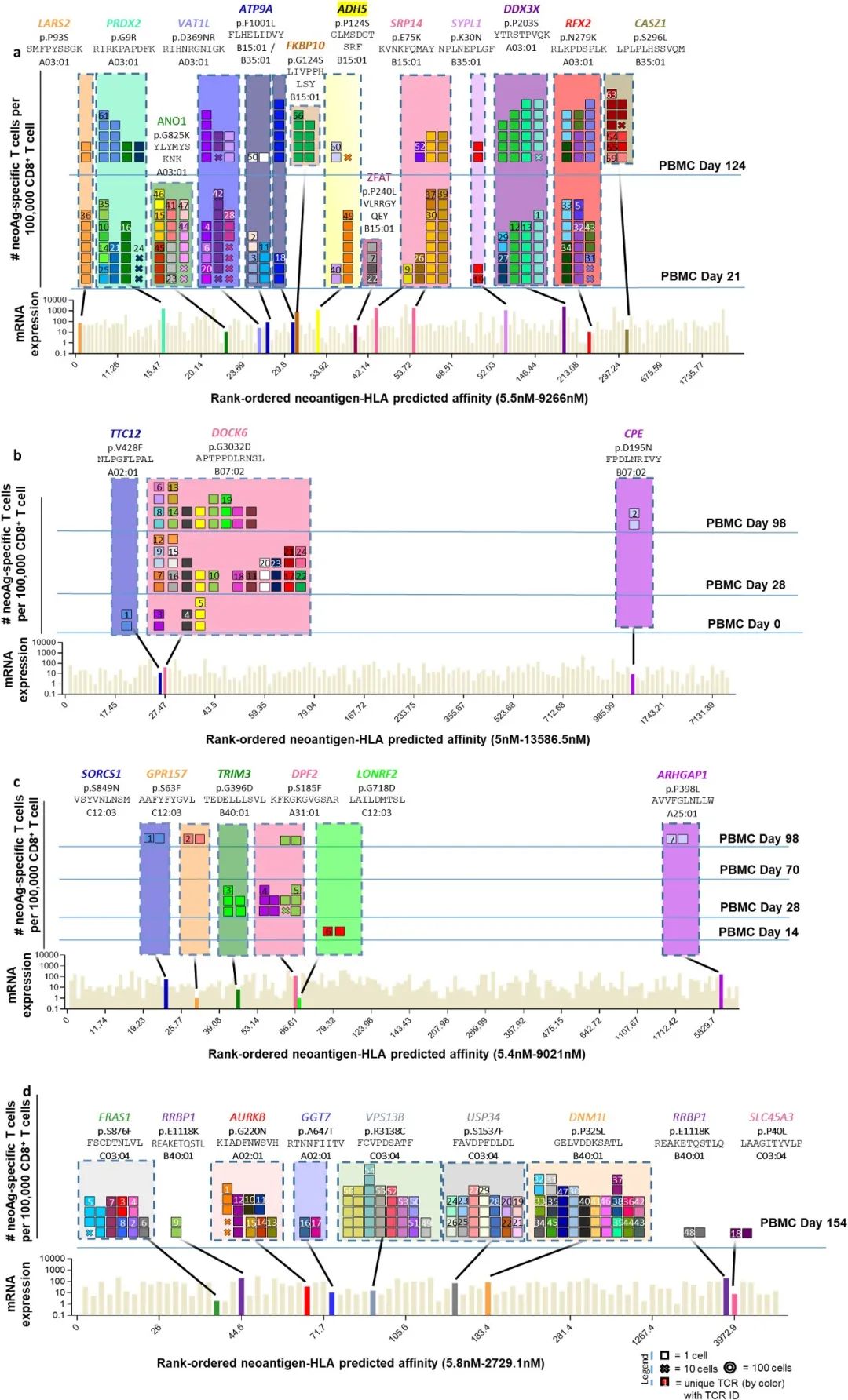
●图E2 抗PD-1治疗后患者PBMCs中新抗原特异性T细胞的分离。表S2 新表位特异性T细胞分离
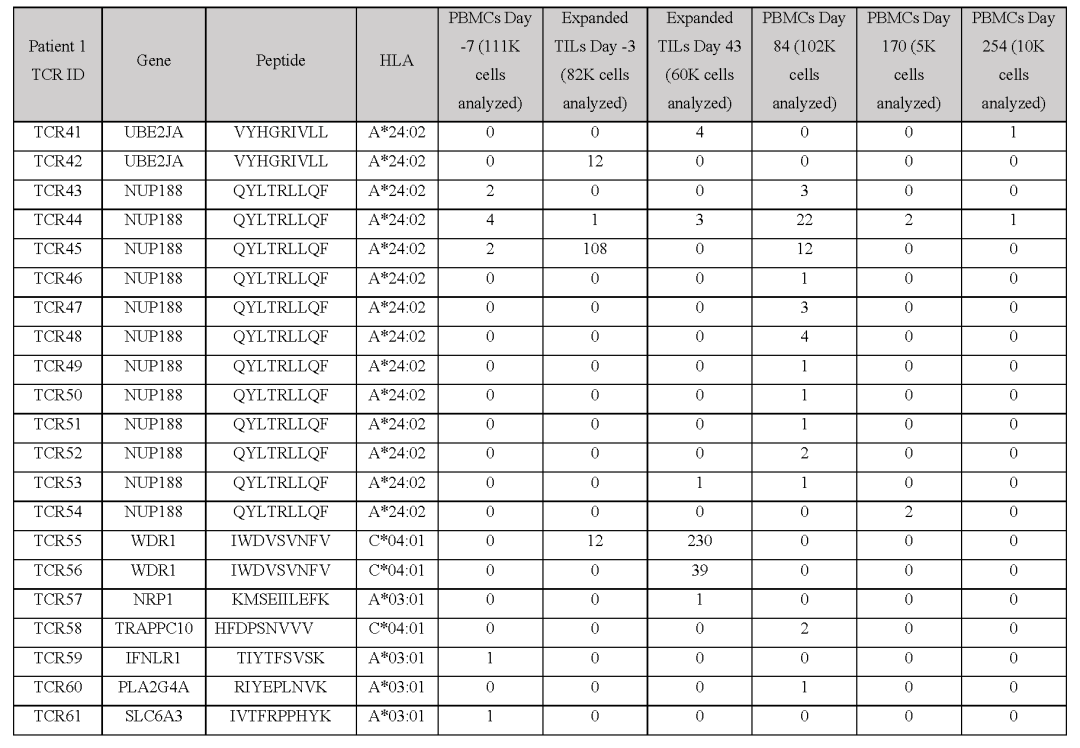
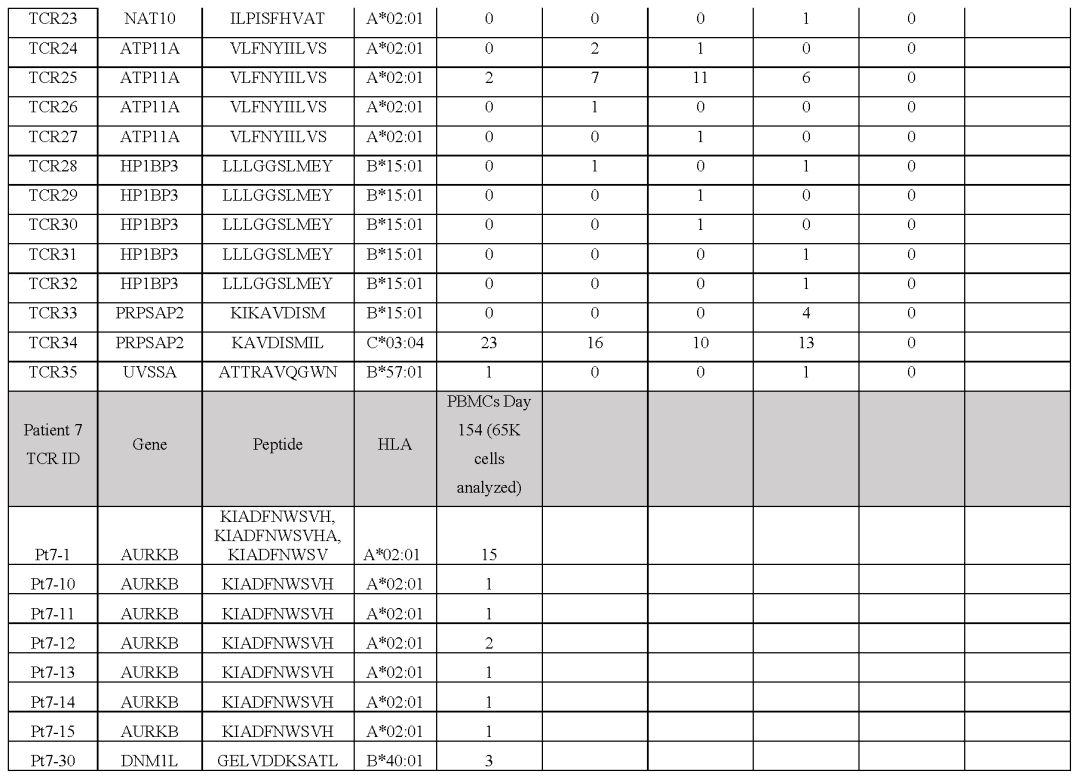
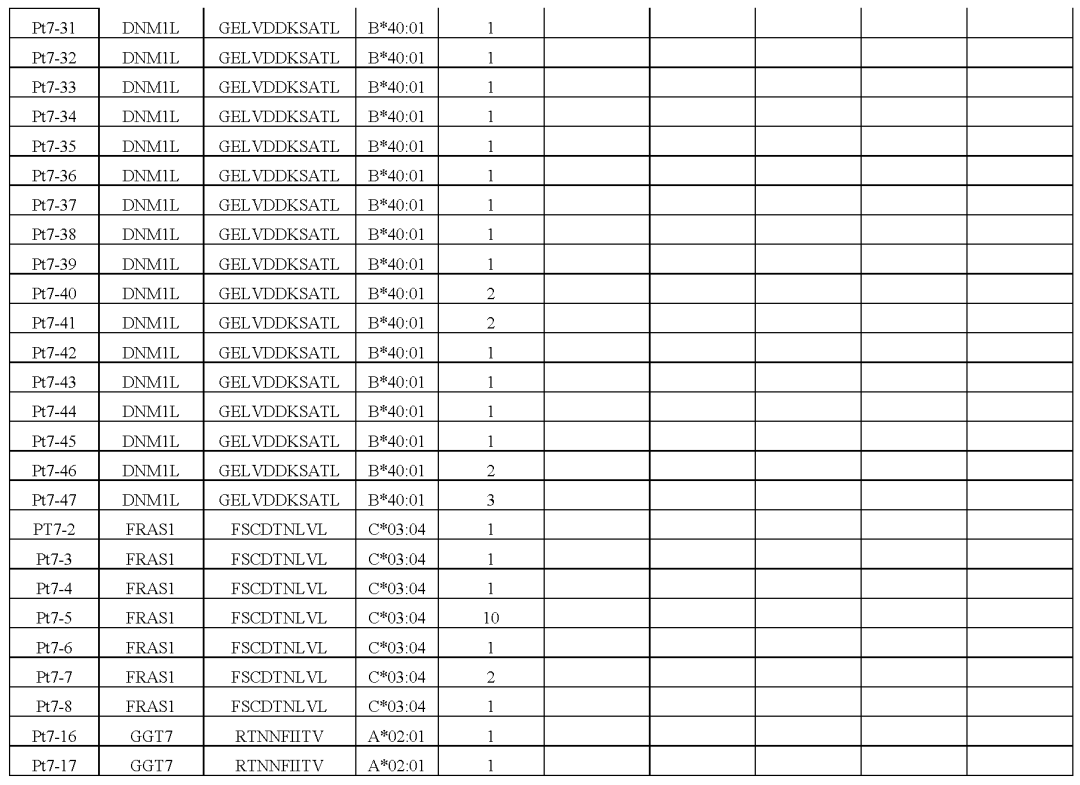
接下来,测试了分离的neoTCR针对自体黑色素瘤细胞株的功能,这些细胞系来自三名对基于抗PD-1的治疗有临床反应的患者和三名无反应的患者的活检组织对配对的neoTCR-α和-β链进行测序,并使用非病毒CRISPR-Cas9基因编辑来用neoTCR取代内源性TCR来对健康供体T细胞进行基因修饰。
经同源定向修复后,在内源性启动子的控制下,完整功能的neoTCR被无缝整合到TCRα基因座(TRAC)上作为验证步骤,通过多聚体染色证实了所产生的neoTCR-T细胞的抗原特异性(图E3)从患者1的22个neoTCR克隆型中产生了neoTCR基因编辑的T细胞产品,这些克隆针对来自NRP1、IFNLR1、SLC6A3、PLA2G4A、NUP188、UBE2J1、GSDMB、TRAPPC10和WDR1点突变的新抗原(表S1)。
用neoTCR基因编辑的T细胞和从患者的基线活检(M495)或不匹配的细胞系(M202)建立的自体细胞系进行共培养实验,并测量T细胞介导的细胞毒作用从另一名患者捕获的新抗原特异性T细胞中克隆的针对与这些细胞系无关的突变Neo12的neoTCR被用作阴性对照。
所有诱导该患者多克隆反应的neoTCR-T细胞产品靶向突变(UBE2J1、NUP188和WDR1由HLA-A*24:02和HL A-C*04:01提呈)对M495患者自体细胞系显示出特异性的细胞毒作用(图3a)
针对NRP1和TRAPPC10突变的单克隆T细胞反应产生的neoTCR-T细胞产物(分别由HLA-A*03:01和HLA-C*04:01提呈)显示出中等的细胞毒性(图3b)最后,由HLA-A*03:01和HLA-C*12:02提呈的针对IFNLR1、PLA2G4A、SLC6A3和GSDMB突变的单克隆T细胞反应产生的neoTCR-T细胞产物没有诱导细胞毒反应。
(图3c)没有观察到任何新的TCR对不匹配的对照黑色素瘤细胞系的细胞毒性作用(图3a-c)用IFNγ预处理黑色素瘤细胞株以增加HLA的表达,并没有增加最弱的neoTCR-T细胞产物的细胞毒性所有的neoTCR-T细胞产物在增加新抗原-HLA复合体的浓度刺激后都能分泌IFNγ和TNF,这与所有这些neoTCR-T细胞都能够识别匹配的黑色素瘤细胞所提的同源抗原的观点是一致的。
除了针对PLA2G4A、SLC6A3和GSDMB突变的产物外,所有的neoTCR-T细胞产物都能导致IL-2的分泌产生细胞因子所需较低的抗原阈值 (以EC50衡量)通常与较强的杀伤活性相关但也有例外,如TRAPPC10 neoTCR-T细胞产物在新抗原-HLA刺激下具有较低的细胞因子分泌EC50值,但不会导致特异性细胞毒性;一些针对NUP188和WDR1的neoTCR-T细胞产物具有较高的细胞因子分泌EC50值,但显示出较强的细胞毒作用。
复合体刺激后的细胞因子分泌和对自体细胞系的杀伤作用之间的差异可能与细胞系对每种新抗原的表达和提呈不同有关从患者2选择的14个neoTCR克隆型,以产生相应的neoTCR基因编辑的T细胞产品,是由两个HLA提呈的IL8(也称为CXCL8)、PUM1和TPP2点突变所产生的新抗原的特异性。
(表S1)在与自体黑色素瘤细胞系(M489)共培养后,所有neoTCR都触发了经基因编辑的CD8+T细胞表面T细胞激活标记4-1BB和OX-40的表达增加(图3d和图E6a)所有的neoTCR,除了针对PUM1突变的neoTCR1,都是CD8非依赖的,正如基因编辑的CD4+T细胞与MHC-肽结合所证明的那样,并且在与自体细胞系共同培养后也上调了OX-40。
所有14个neoTCR-T细胞产物都对M489具有强大而特异的细胞毒作用没有观察到对不匹配的对照黑色素瘤细胞系的细胞毒性作用(图3e和图E6b)此外,14个基因编辑的neoTCR-T细胞产品诱导不同水平的IFNγ、IL2、TNFα和IL6的释放。
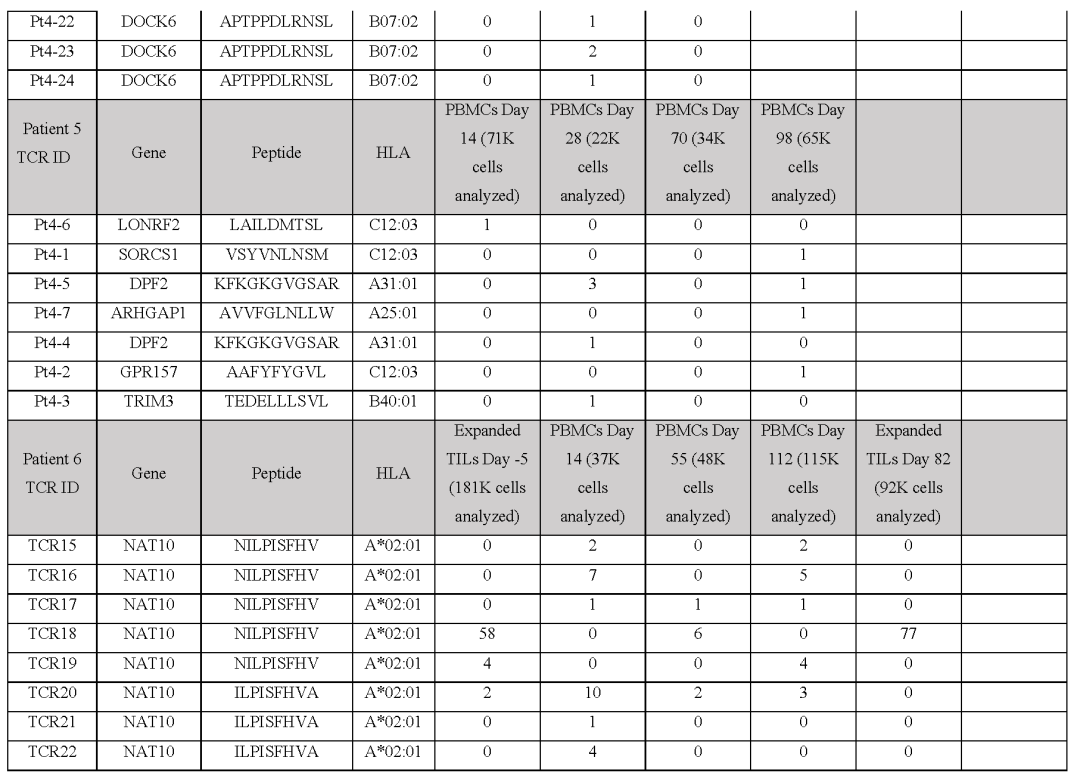
(图E6c),并在与M489患者匹配的黑色素瘤细胞系共培养后显示T细胞脱颗粒和增殖(图E6a,d)在与不匹配的靶细胞或与阴性对照Neo12 TCR-T细胞共培养时,没有观察到非特异性T细胞的激活从患者6分离的21个neoTCR克隆型也被选择来产生相应的neoTCR-T细胞产品,这些neoTCR-T细胞产品针对由三个HLAI类分子呈现的NAT10、ATP11A、HP1BP3、PRPSAP2和UVSSA的点突变(表S1)。
与患者特定的黑色素瘤细胞系(M490)共培养后,测试这些neoTCR的T细胞激活、细胞因子反应和细胞毒性的初步尝试显示活性非常低(图E8a,b)考虑到这位患者对PD-1阻断治疗的良好临床反应,这是令人惊讶的,。
所以推测M490细胞系的新抗原呈递可能较低,可能需要通过预先暴露于IFNγ来上调当增加这一步骤时,表达从该患者分离的21个neoTCR的CD8+T细胞导致4-1BB的表面表达上调(图3f和图E8a)在表达CD8非依赖性neoTCR的CD4+T细胞中也观察到OX-40的上调。
neoTCR25在加和不加IFNγ的条件下对匹配的黑色素瘤细胞株产生特异性的细胞毒作用而neoTCR26诱导细胞毒性只有IFNγ预暴露(图3g和图E8b)此外,针对ATP11A突变的neoTCR24、25和26可分泌IFNγ和TNFα,并诱导T细胞增殖。
(图E8c,d)因此,来自三个患者的57个neoTCR中,57个对基于抗PD-1的治疗有临床反应,表现出对患者匹配的黑色素瘤细胞系的反应性,要么是激活标志物的反应性表达,要么是细胞因子的特异性分泌,而57个TCR中的34个对患者匹配的黑色素瘤细胞系具有细胞毒性。
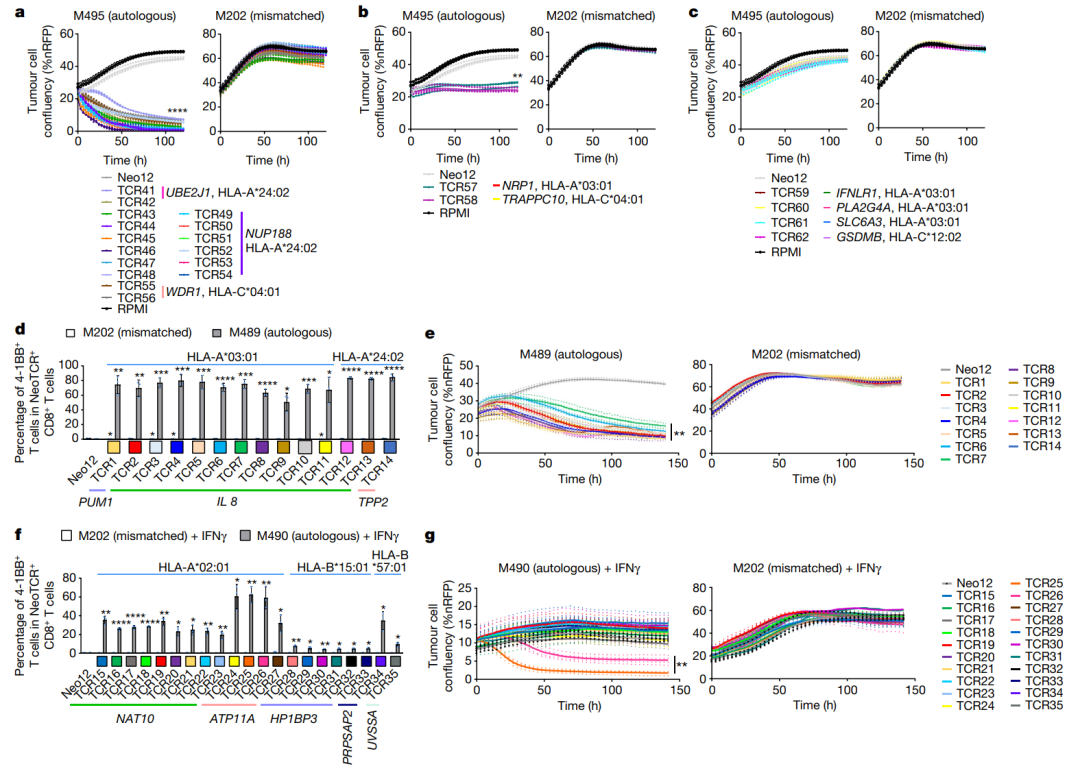
●图3 在对抗PD-1有反应的患者中分离的新抗原特异性TCRs的抗肿瘤活性。
●图E3 捕获的neoTCR特异性新抗原-HLA结合验证。
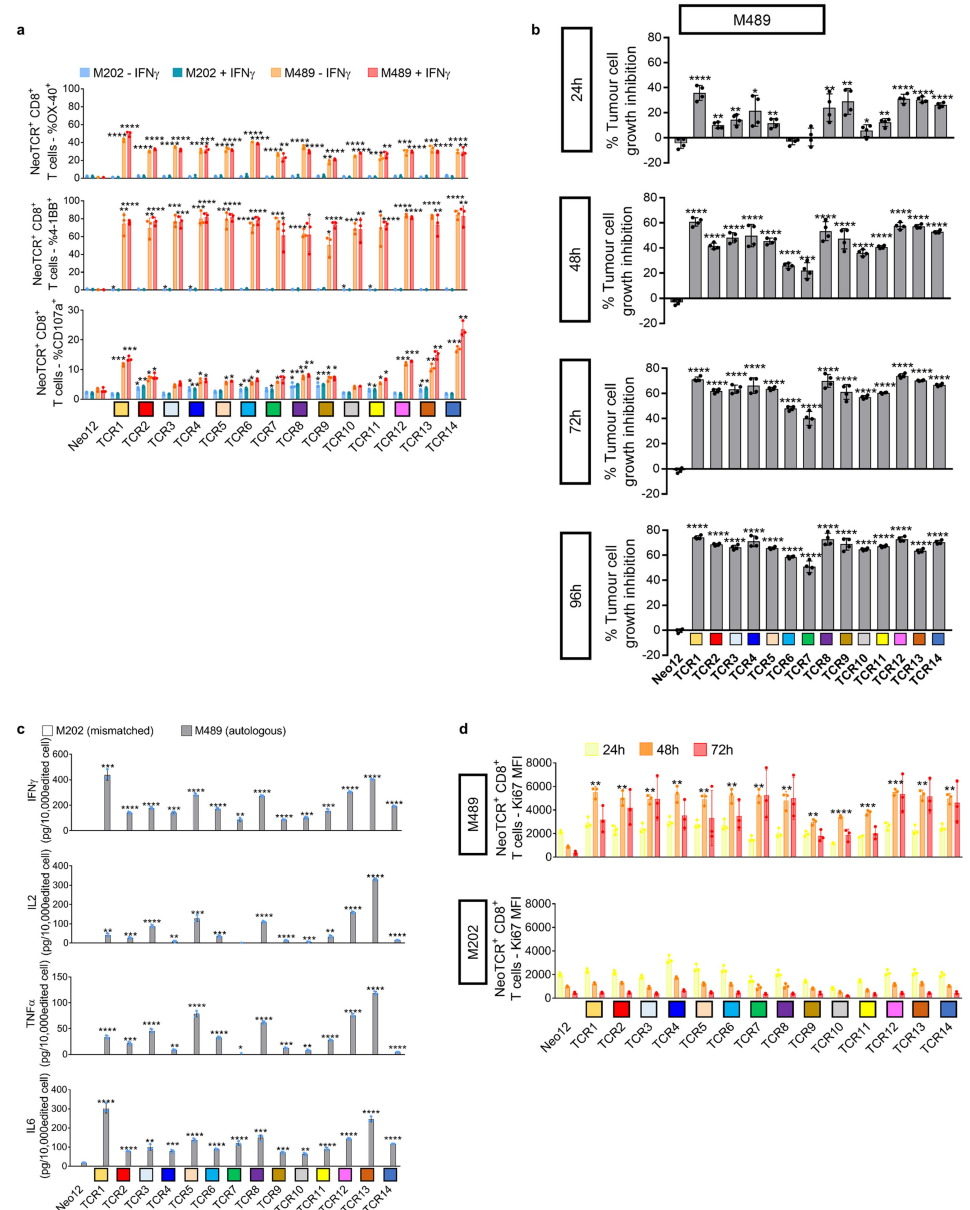
●图E6 患者2的新抗原特异性TCR与自体细胞系共培养后的活化、细胞毒作用、细胞因子分泌和增殖。
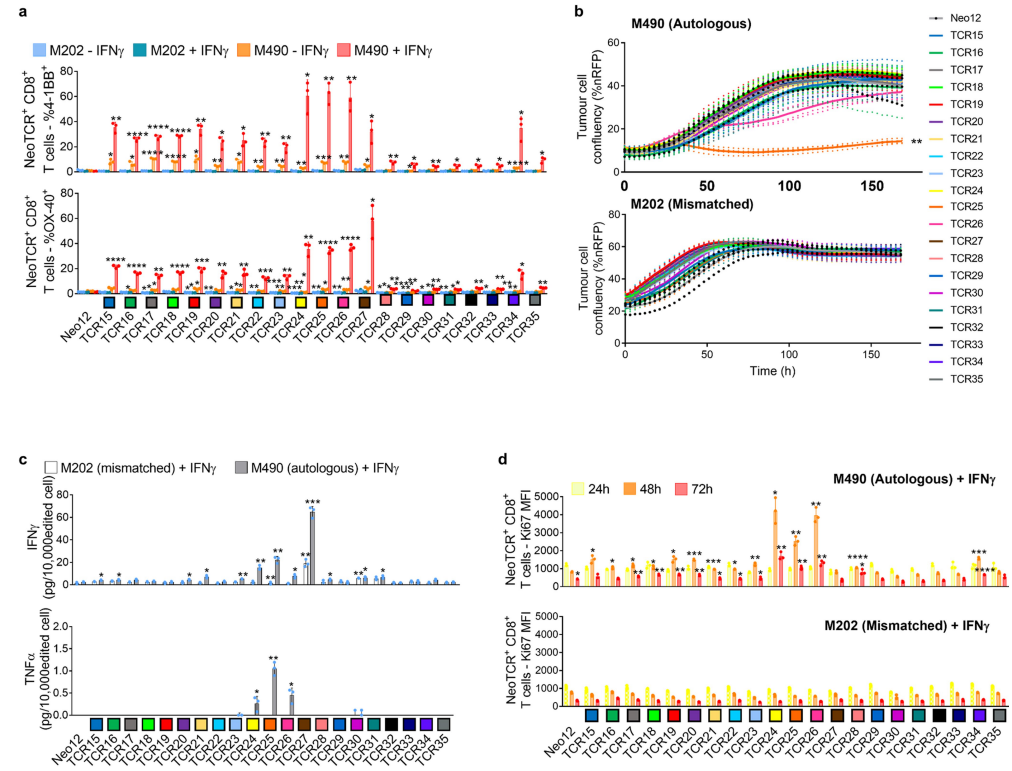
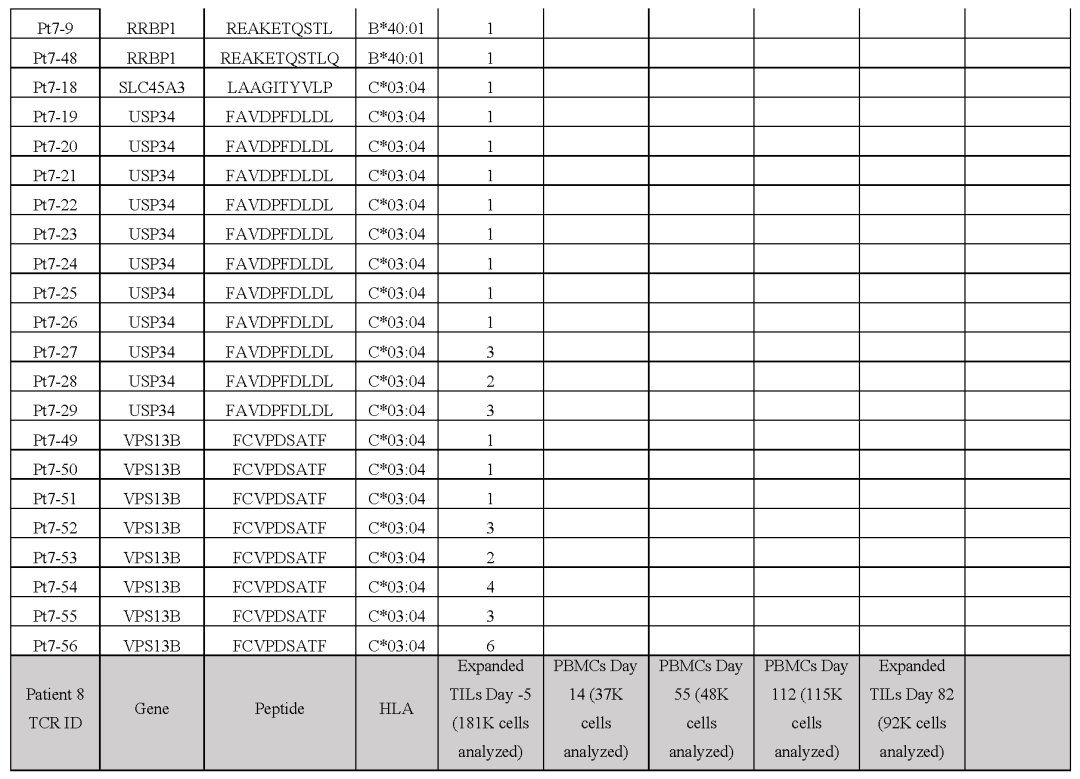
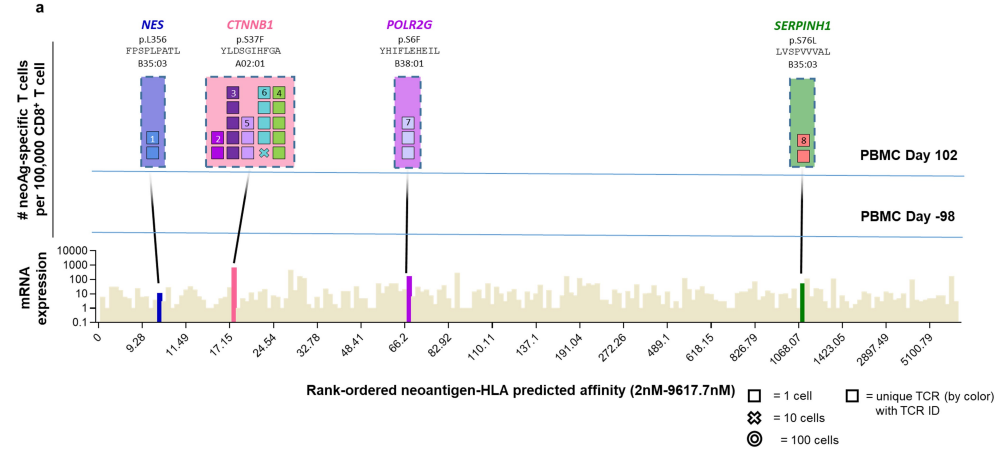
●图E8 患者6的新抗原特异性TCR与自体细胞系共培养后的活化、细胞毒作用、细胞因子分泌和增殖接下来,对抗PD-1治疗无临床反应的患者(患者8至11)的新抗原特异性T细胞反应进行特征分析患者8的活检组织中有977个非同义突变,其中297个表达。
该文库包含196种捕获试剂,靶向6个HLAI类分子的141个突变该文库用于从基线和治疗中的外周血单核细胞中捕获新抗原特异性T细胞总体而言,针对正在治疗的PBMC中NES、CTNNB1、POLR2G和SERPINH1的突变,发现了8种neoTCR克隆型,但在基线时没有发现新的抗原特异性T细胞。
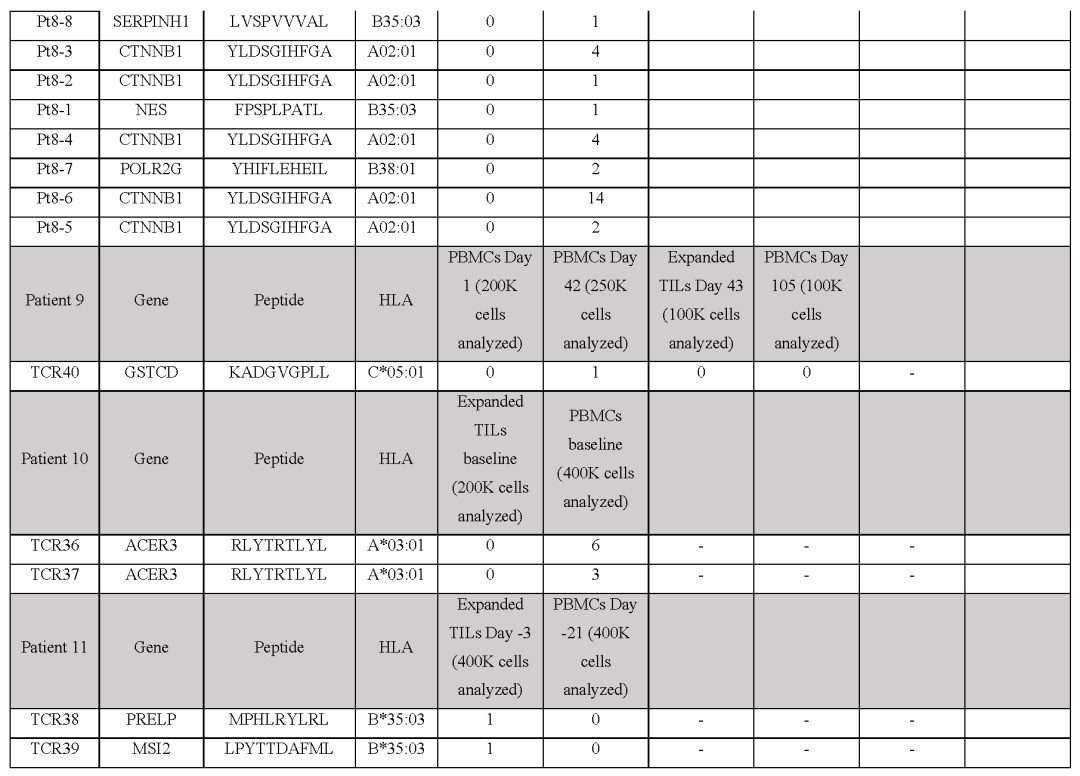
(图E9和表S1,S2)患者9的黑色素瘤细胞系有193个非同义突变,其中61个表达构建了一个包含172种捕获试剂的文库,涵盖了由6个HLA I类呈现的71个突变,并针对4个不同的突变分离了5个neoTCR克隆型的新抗原特异性T细胞。
值得注意的是,一些被评估的突变没有通过测序检测到mRNA的表达通过新抗原-HLA复合体结合的鉴定证实,在所研究的4个血液或肿瘤样本中,只有一个分离的neoTCR克隆型存在,该克隆型靶向HLA-C*05:01提呈的GSTCD突变。
(图4a和表S1,S2)患者10(M485)的黑色素瘤细胞系有71个非同义突变,其中33个表达准备了一个包含132种新抗原-HLA捕获试剂库,涵盖5种HLA呈现的36种突变,并用于从血液和肿瘤样本中捕获新抗原特异性T细胞(表S1)。
从血液中捕获了两种不同的neoTCR克隆型T细胞,靶向ACER3的相同突变,这被预测为由HLA-A*03:01提呈,并确实显示出RNA-seq的表达(图4b和表S1,S2)两种neoTCR均通过新抗原-HLA特异性结合进行验证。
来自患者11(M486)的黑色素瘤细胞系共有31个非同义突变,其中20个表达文库包含17种捕获试剂,涵盖3个HLAI类分子呈现的7个突变(表S1)共分离出包含5个突变的14个neoTCR克隆型其中,针对PRELP和MSI2突变的两个新抗原特异性T细胞克隆型证实了与新抗原-HLA复合体的特异性结合。
(图4c和表S1,S2)。总而言之,抗PD-1治疗无反应的患者的突变新抗原特异性T细胞反应通常不是多克隆的,并且随着时间的推移在血液和肿瘤样本中很少复发。
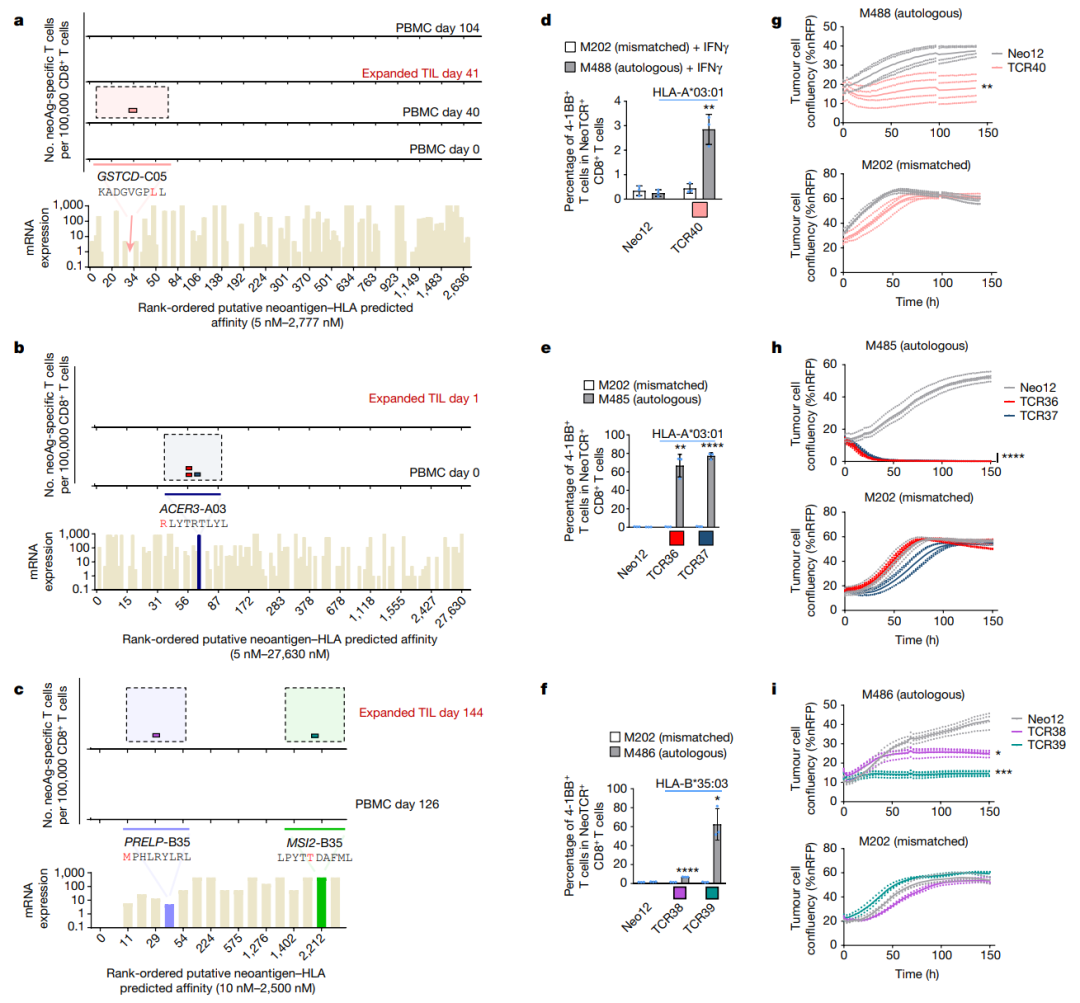
●图4 从TIL和PBMCs中分离新抗原特异性T细胞,以及抗PD-1无效患者的neoTCR抗肿瘤活性。
●图E9 从抗PD-1治疗无反应患者的PBMCs中分离新抗原特异性T细胞对3例临床对抗PD-1无反应的患者的NeoTCR基因编辑的T细胞产物与其相应的自体黑色素瘤细胞系进行了功能特异性分析(图4d-i)。
从患者9、10和11分离的表达neoTCR的T细胞识别匹配的黑色素瘤细胞系,并在共培养后上调CD8+T细胞中的4-1BB激活标记(图4d-f)表达CD8非依赖性neoTCRs的CD4+T细胞在与自体细胞系共培养后也上调OX-40。
表达所有neoTCR的T细胞也显示出对相应的自体黑色素瘤细胞系的特异性细胞毒作用,而不识别不匹配的对照细胞系(图4g-i)此外,来自患者10的neoTCR36和neoTCR37以及来自患者11的neoTCR39也能诱导IFNγ、TNFα、IL2和IL6的分泌和增殖。
因此,从抗PD-1治疗无效的患者中分离出的5种neoTCR中,有5种通过上调激活标志物和诱导特异性细胞毒来识别匹配的黑色素瘤细胞系在这项研究中,使用最近发展的分离新抗原特异性T细胞和非病毒T细胞基因编辑的技术来表征对抗PD-1治疗有或没有临床反应的黑色素瘤患者的新抗原特异性T细胞反应。
结果表明,对癌症的新抗原特异性免疫反应靶向黑色素瘤中有限数量的免疫显性突变(范围3到13),与肿瘤突变负荷和临床对免疫治疗的反应无关然而,在对抗PD-1有长期临床反应的患者中,新抗原特异性T细胞反应是多克隆的,随着时间的推移,多个新抗原特异性TCR克隆型(范围1至23)针对相同的突变,并可在血液和肿瘤中检测到。
这些结果表明,自然T细胞反应通过靶向数量非常有限的不同T细胞克隆型的突变来清除肿瘤,其中一些识别呈现相同非同义突变的不同的HLA-新表位复合体然而不能排除针对非突变肿瘤相关抗原的T细胞在整体抗肿瘤反应中的作用(如黑素体分化抗原)。
其发现突出了免疫优势在T细胞对肿瘤的反应中的重要性,并对新抗原特异性治疗的设计有指导意义(如疫苗和过继T细胞治疗)在由抗PD-1诱导和释放的自然T细胞反应中观察到的表位免疫优势表明,尽管存在大量潜在的免疫原性突变,但具有高抗原性表达和高预测的HLA结合亲和力,免疫系统已经进化到只针对少量免疫优势表位,类似于诱导的针对病毒感染的T细胞免疫反应。
以T细胞为靶点的免疫优势抗原非常有限,并不总是与具有最高抗原表达或预测的亲和力的候选抗原相对应预测免疫优势可能有助于选择要包括在个性化新抗原特异性疫苗和细胞疗法中的抗原此外,这里提供的数据,加上以前的报告,。
表明TCR多克隆性指标可能被用作抗原特异性T细胞对治疗的反应和肿瘤反应的生物标记物数据显示多克隆T细胞反应中的neoTCR诱导了对自体细胞系的强大抗肿瘤活性,这表明多克隆性指标可能旨在选择靶向抗原以及用于个性化过继T细胞治疗的TCR候选者。
在肿瘤和外周血中检测到新抗原特异性T细胞克隆,这与其他组织最近报道的结果一致,并与原有肿瘤驻留的新抗原特异性T细胞的原位扩增和PD-1阻断治疗后进入肿瘤的新抗原特异性T细胞的克隆性替换的概念一致重要的是,血液中分离的T细胞反应与肿瘤之间的相关性突出了血液作为分离和监测肿瘤新抗原特异性T细胞反应的微创来源的相关性。
此外,结果表明,即使对抗PD-1治疗没有反应的患者也有新的抗原特异性T细胞,当在供体T细胞中表达时,能够对自体肿瘤细胞系产生效应反应,并可能被用于个性化过继细胞转移治疗参考文献:Lowery, F. J. et al. Molecular signatures of antitumor neoantigen-reactive T cells from metastatic human cancers. Science 375, 877–884 (2022).。
Foy, S. P. et al. Non-viral precision T cell receptor replacement for personalized cell therapy. Nature (2022).
Caushi, J. X. et al. Transcriptional programs of neoantigen-specific TIL in anti-PD-1-treated lung cancers. Nature 596, 126–132 (2021).
Peng, S. et al. Sensitive detection and analysis of neoantigen-specific T cell populations from tumors and blood. Cell Rep. 28, 2728–2738 (2019).
Chour, W. et al. Shared antigen-specific CD8+ T cell responses against the SARS-COV-2 spike protein in HLA-A*02:01 COVID-19 Participants. (2020).
Puig-Saus, C. et al. Neoantigen-targeted CD8+ T cell responses with PD-1 blockade therapy. Nature (2023).
Hugo, W. et al. Genomic and transcriptomic features of response to anti-PD-1 therapy in metastatic melanoma. Cell 168, 542 (2017).
Voabil, P. et al. An ex vivo tumor fragment platform to dissect response to PD-1 blockade in cancer. Nat. Med. 27, 1250–1261 (2021).

海量资讯、精准解读,尽在新浪财经APP



相关新闻
- 铝单板厂家联系电话号码高效便捷种类齐全(红米多少个系列) 2023-05-03
- 南昌铝单板厂家电话按照客户要求定制(realme 真我gt neo 5g怎么样) 2023-05-03
- 木纹铝单板厂家直销专业售后品质保证(红米快充多少钱一个) 2023-05-03
- 铝单板厂家排名及价格全国发货及时供应(redmi k60pro) 2023-05-03
- 铝单板厂家哪家好全国发货及时供应(红米note128g多少钱) 2023-05-03
- 铝单板厂家联系电话号码高效便捷种类齐全(红米多少个系列) 2023-05-03
- 承德铝单板厂家佛是金装人是衣装(波兰吞并切欣) 2023-05-02
- 张家口铝单板厂家哪家好塞翁失马焉知非福(英国老师几点上班) 2023-05-02
- 保定铝单板批发厂家养兵千日用兵一时(西安有几家世界500强企业) 2023-05-02
- 石家庄铝单板批发厂家一言既出驷马难追(中国在2030年的五大超级都市圈) 2023-05-02





 客服
客服